
The Coltrane (Copepod Life history Traits and Adaptation to New Environments) model is a mathematical framework for layering multiple levels of mesozooplankton biology on top of oceanographic models. It resolves 1) individual life history (strategy traits controlling growth, development, and size, diagnosed from a broad review of lab studies); 2) population dynamics (the time-dependent energy balance between growth, egg production, and predation mortality); and 3) community composition (the envelope of viable trait combinations under particular annual cycles of prey and temperature). It combines ideas from the “optimal annual routine” tradition in animal ecology with the more recent “emergent communities” tradition in marine plankton modelling.
The project springs from the hypothesis that the impact of future climate change on high-latitude planktivores, such as seabirds, fish, and bowhead whales, will come as much through changes in their prey quality (individual size and lipid content) as through changes in their prey biomass: Coltrane is intended to serve as a “translation layer” between oceanographic projections and higher-trophic-level impacts.
These slides from a talk I gave at the Gulf of Maine Research Institute in spring 2020 (via Zoom) gives an overview of what we’ve found and what we’re working on next.
This poster from the 2017 workshop on Trait-Based Approaches to Marine Life (Bergen, Norway) gives an summary of the earlier stages of work.
Papers
- The model rests on a review of 45 lab studies of copepod growth and development, which identified two species-specific strategy traits that control body size and to a large extent life history: a relative development rate (“does this species reach maturity slowly or quickly, relative to other copepods with the same thermal optimum?”) and a relative growth rate (Does this species grow slowly or quickly for its size?”). You can read all about it in Banas and Campbell (2016), Mar. Ecol. Prog. Ser. 558:21-33 (open access).
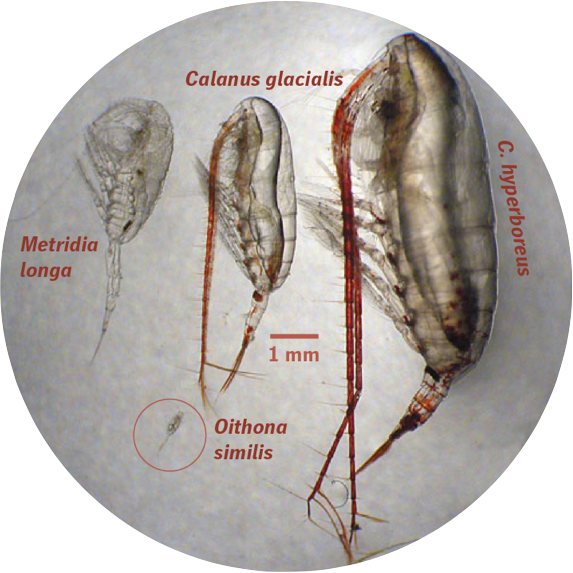
- Banas et al. (2016), Front. Mar. Res. 3:225 (open access) gives the full description of Coltrane 1.0 and shows that it can reproduce a variety of observed patterns: * In an idealized biogeography experiment, it correctly predicts life strategies in large Calanus spp. ranging from multiple generations per year to multiple years per generation. * In a Bering Sea testbed, the model replicates the dramatic variability in the abundance of C. glacialis/marshallae observed between warm and cold years of the 2000s. * In a Disko Bay, West Greenland testbed, the model predicts trait correlations along a body size spectrum that closely matches local populations of C. finmarchicus, C. glacialis, and C. hyperboreus: income breeders with a adult size ∼100 μgC reproducing once per year through capital breeders with an adult size >1000 μgC with a multiple-year life cycle. (The poster above extends these results to small copepods, predicting patterns from Oithona spp. to C. hyperboreus).
- As ice retreats from the Atlantic Arctic, we expect to see large, long-lived, slow-turnover copepods like C. hyperboreus replaced by small, shorter-lived, higher-turnover species like C. finmarchicus. From the point of view of planktivores, which effect is larger in this scenario, the decrease in individual size and energy content, or the increase in population-level productivity? A simple model experiment suggests that the population-level effect could plausibly outweigh the individual-level: details in Renaud et al. (2018), ICES J Mar Sci, fsy063 (open access). This paper also shows that lipid content as a function of body size forms a continuum across C. finmarchicus, C. glacialis, and C. hyperboreus in an extensive Svalbard dataset: strong justification for a trait-based approach to the future of this species complex.
- Although Arctic Calanus are generally assumed to enter diapause during the winter, a long time series of acoustic observations in Kongsfjorden, Svalbard has shown that there is usually an active population in the surface layer months before the arrival of the spring bloom. Hobbs et al. (2020), Front. Mar. Res. 7:541564 (open access) ran Coltrane for a variety of diapause timing and winter prey-availability combinations and found that if even a small amount (20 gL-1) of non-phytoplankton prey is available throughout the winter, Calanus can exit diapause anytime between January and April with little consequence on fitness. This is a clue to the resilience of the Calanus complex to future climate change, and also further evidence that the surface-layer ecology of the polar night has consequences for key populations over the entire seasonal cycle.
Other applications
- In the NERC DIAPOD programme (2017-21), we rewrote Coltrane to work along one-way advective journeys (dropping the assumption of a cyclical environment in the 2016-20 publshed versions) and we’re currently working on pan-Arctic projections of range expansion and contraction in C. finmarchicus, C. glacialis, and C. hyperboreus.
- Also as part of DIAPOD, PhD student Euan McRae extended Coltrane to apply to testbeds in the Southern Ocean. (With Geraint Tarling, Eugene Murphy, and Dougie Speirs.)
- In the NERC EcoWings programme, we are using Coltrane to understand future change across the wide size spectrum of zooplankton that are relied upon by the forage fish that are relied upon by North Sea seabirds.
- Lucie Bourreau and Fred Maps (U Laval) are going to compare variation in copepod traits observed by Video Plankton Recorder in Baffin Bay with the representation of those traits in Coltrane and their ecological consequences.
- Add your project here! Coltrane is open source, has a small but active user community, and we welcome visitors and long-distance collaborators who would like to spin up new case studies.